
Diphyllobothriasis
[Dibothriocephalus latus (=Diphyllobothrium latum)] [Dibothriocephalus spp.][Adenocephalus pacificus]
Causal Agents
Several members of the cestode (tapeworm) family Diphyllobothriidae are known to infect humans. These pseudophyllidean cestodes have a scolex bearing bothria (grooves), instead of suckers as in the cyclophyllidean cestodes (the group including nearly all human-infecting species). All species associated with human diphyllobothriid infections have marine or aquatic life cycles and transmission occurs via ingestion of undercooked fish.
Recent research incorporating morphologic and molecular data has led to the re-classification and re-naming of most of the human-infecting diphyllobothriids. Dibothriocephalus latus (=Diphyllobothrium latum), the “broad fish tapeworm”, is usually assumed to be the most common agent of human diphyllobothriasis. However, it is possible that many historical cases were falsely attributed to this species. Dibothriocephalus nihonkaiense (=Diphyllobothrium nihonkaiense), Dibothriocephalus dendriticus (=Diphyllobothrium dendriticum), Diphyllobothrium stemmacephalum (=Diphyllobothrium stemmacephalum; =Diphyllobothrium yonagoense), Diphyllobothrium balaenopterae (=Diplogonoporus grandis; =Diplogonoporus balaenoptera), and Adenocephalus pacificus (=Diphyllobothrium pacificum) are also known to infect humans. Sporadic case reports exist involving several other diphyllobothriid species, although some of the species identifications in these reports are of questionable validity.
Life Cycle
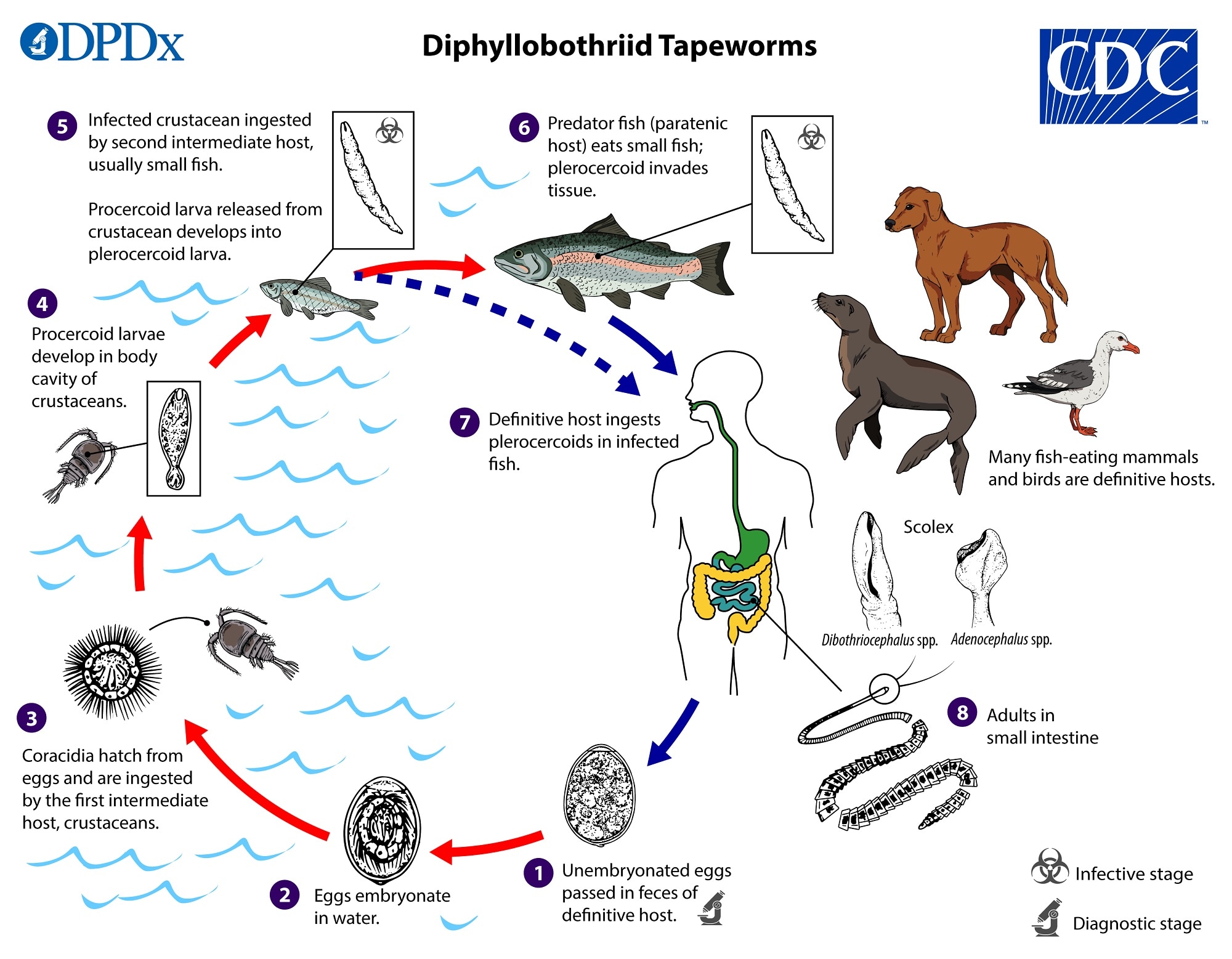
Eggs are passed unembryonated in feces  . Under appropriate conditions, the eggs mature (approximately 18 to 20 days)
. Under appropriate conditions, the eggs mature (approximately 18 to 20 days)  and yield oncospheres which develop into a coracidia
and yield oncospheres which develop into a coracidia  . After ingestion by a suitable crustacean (first intermediate host) the coracidia develop into procercoid larvae
. After ingestion by a suitable crustacean (first intermediate host) the coracidia develop into procercoid larvae  . Procercoid larvae are released from the crustacean upon predation by the second intermediate host (usually a small fish) and migrate into the deeper tissues where they develop into a plerocercoid larvae (spargana), which is the infectious stage for the definitive host
. Procercoid larvae are released from the crustacean upon predation by the second intermediate host (usually a small fish) and migrate into the deeper tissues where they develop into a plerocercoid larvae (spargana), which is the infectious stage for the definitive host  . Because humans do not generally eat these small fish species raw, the second intermediate host probably does not represent an important source of human infection. However, these small second intermediate hosts can be eaten by larger predator species that then serve as paratenic hosts
. Because humans do not generally eat these small fish species raw, the second intermediate host probably does not represent an important source of human infection. However, these small second intermediate hosts can be eaten by larger predator species that then serve as paratenic hosts  . In this case, the plerocercoid migrates to the musculature of the larger predator fish; humans (and other definitive host species) acquire the parasite via consumption of undercooked paratenic host fish
. In this case, the plerocercoid migrates to the musculature of the larger predator fish; humans (and other definitive host species) acquire the parasite via consumption of undercooked paratenic host fish  . In the definitive host, the plerocercoid develops into adult tapeworms in the small intestine. Adult diphyllobothriids attach to the intestinal mucosa by means of two bilateral groves (bothria) of their scolex
. In the definitive host, the plerocercoid develops into adult tapeworms in the small intestine. Adult diphyllobothriids attach to the intestinal mucosa by means of two bilateral groves (bothria) of their scolex  . The adults can reach more than 10 m in length, with more than 3,000 proglottids. Immature eggs are discharged from the proglottids (up to 1,000,000 eggs per day per worm) and are passed in the feces. Eggs appear in the feces 5 to 6 weeks after infection.
. The adults can reach more than 10 m in length, with more than 3,000 proglottids. Immature eggs are discharged from the proglottids (up to 1,000,000 eggs per day per worm) and are passed in the feces. Eggs appear in the feces 5 to 6 weeks after infection.
Hosts
Intermediate and paratenic hosts include a wide diversity of freshwater and marine fishes, commonly perch and pike for Dibothriocephalus latus and salmon for D. nihonkaiense.
Definitive host specificity among the medically important diphyllobothriids is low. In addition to humans, other carnivorous, fish-eating mammals and a few birds can also serve as definitive hosts. Non-human definitive hosts for D. latus, D. nihonkaiense, and D. dendriticus include a variety of carnivorous mammals (canids, felids, bears, and mustelids); of these, only D. dendriticus is known to parasitize birds, particularly gulls. Pinnipeds are the natural definitive host for Adenocephalus pacificus and it has also been found in dogs and jackals. Among the cetacean-associated species, Diphyllobothrium stemmacephalum is found mostly in dolphins and porpoises, and D. balaenopterae in whales.
Geographic Distribution
Dibothriocephalus latus has a relatively wide distribution, but is most common in circumpolar regions; it is also known to occur very rarely in South America, perhaps through importation and aquaculture. D. nihonkaiense is endemic to the northern Pacific Ocean and cases are common in East Asian countries where raw fish dishes are popular. D. dendriticus has a generally holarctic distribution and is most prevalent in the arctic. While found in hosts across the Pacific Ocean, a large majority of Adenocephalus pacificus cases occur on the Pacific coast of South America and one case has occurred in South Australia. Diphyllobothrium stemmacephalum and D. balaenopterae are considered to have a cosmopolitan marine distribution.
Note that the global trade and consumption of wild-caught fish leads to human cases occurring regularly outside of naturally endemic regions.
Clinical Presentation
Diphyllobothriasis can be a long-lasting infection, up to 25 years. Most infections are asymptomatic but gastrointestinal symptoms may occur in some patients. Aberrant migration of proglottids can cause cholecystitis or cholangitis. Rarely, massive infections may cause intestinal obstruction.
Diphyllobothriid eggs
Diphyllobothriid eggs are oval or ellipsoidal and range in size from 55 to 75 µm by 40 to 50 µm. There is an operculum at one end that can be inconspicuous, and at the opposite (abopercular) end is a small knob that can be barely discernible. The eggs are passed in the stool unembryonated. Although eggs of Adenocephalus pacificus are generally smaller than those of Dibothriocephalus spp. and Diphyllobothrium spp., there is a degree of overlap and thus species identification based on eggs may not be reliable.










Mature diphyllobothriids are large tapeworms reaching 2—15 meters in length, with occasional larger specimens. The scolex always has two bothria (grooves). The general size and shape of the scolex may be subject to intraspecific variability, though some species-level differences have been described historically. Proglottids are broader than long, with a single genital pore that opens in the middle of the ventral surface; fully mature specimens may be comprised of a 2,000—5,000 proglottids. The ovaries are characteristically rosette-shaped.











Laboratory Diagnosis
Microscopic identification of eggs in the stool is the basis of family level diagnosis; genus level identification based on eggs is difficult due to overlap in morphological features. Eggs are usually numerous and can be demonstrated without concentration techniques. Identification of proglottids passed in the stool is also of diagnostic value.
PCR on DNA extracted from concentrated eggs or proglottids (either fresh or preserved with a PCR-compatible fixative) is usually necessary for species-level identification, as clinical samples usually lack sufficient morphologically-relevant material from adult worms (e.g. scolex, neck, strobila). Also, treatment may distort morphologic features of expelled cestodes tissues. Molecular identification is generally only used in research capacities at present.
More on: Morphologic comparison with other intestinal parasites.
Laboratory Safety
Standard precautions for the processing of stool samples apply. Unlike taeniid tapeworm eggs (e.g. Taenia, Echinococcus, Hymenolepis), diphyllobothriid eggs are not directly infectious to humans.
Suggested Reading
Ikuno, H., Akao, S. and Yamasaki, H., 2018. Epidemiology of Diphyllobothrium nihonkaiense Diphyllobothriasis, Japan, 2001–2016. Emerging Infectious Diseases, 24(8), p.1428.
Waeschenbach, A., Brabec, J., Scholz, T., Littlewood, D.T.J. and Kuchta, R., 2017. The catholic taste of broad tapeworms–multiple routes to human infection. International Journal for Parasitology, 47 (13), pp.831-843.
Scholz, T. and Kuchta, R., 2016. Fish-borne, zoonotic cestodes (Diphyllobothrium and relatives) in cold climates: a never-ending story of neglected and (re)-emergent parasites. Food and Waterborne Parasitology, 4, pp.23-38.
Kuchta, R., Serrano-Martínez, M.E. and Scholz, T., 2015. Pacific broad tapeworm Adenocephalus pacificus as a causative agent of globally reemerging diphyllobothriosis. Emerging infectious diseases, 21 (10), p.1697.
DPDx is an educational resource designed for health professionals and laboratory scientists. For an overview including prevention, control, and treatment visit www.cdc.gov/parasites/.